Marina Arce
Introducción
Los términos informacionales se han vuelto ubicuos en la biología. Frases como “información genética”, “código genético”, decir que cierto gen “codifica para” cierta proteína, “DNA codificante” (1), o “RNA mensajero” son prácticamente ineludibles al revisar la literatura. En 1958, Francis Crick (2) formuló el Dogma central afirmando que “una vez que la información pasa a la proteína ya no puede volver a salir” (Kay, 2000). Más recientemente, en la edición 2017 del libro de texto Evolution, de Douglas Futuyma se lee: “La aparente teleología (3) es causada por la operación de un programa—información codificada o preestablecida que reside en las secuencias de DNA” (Futuyma, 2017). Citando a Paul Griffiths:
«La concepción de que el desarrollo (4) de las características es la expresión de información acumulada en el genoma (5) durante la evolución y que la herencia es la transmisión de esta información de una generación a la siguiente, le parecerá a la mayoría de los biólogos como sentido común (Griffiths, 2017).»
Como puede observarse en estas citas, la información es conceptualizada como un programa interno del desarrollo que se encuentra en el DNA. Se trata de un programa que especifica, codifica, desde adentro al organismo previo a su existencia en el mundo. Es la tesis de este trabajo que la metáfora informacional respalda una falsa dicotomía entre organismo y ambiente, en la cual los genes, como repositorio informacional, ya contienen la especificación del organismo de manera descontextualizada y aislada del ambiente.
Método
Para ello busco emplear la historia para desnaturalizar la cualidad informacional del DNA, y con ello la visión asimétrica que coloca a los genes aparte y por encima de los demás factores y procesos del desarrollo. Para ello me baso en la idea que Joan Wallach Scott retoma de Michel Foucault referente a hacer una historia del presente, “una historia que ‘sirva para mostrar cómo lo-que-es no siempre ha sido’ y mostrar ‘por qué y cómo lo-que-es puede dejar de serlo” (Scott, 2009). Y también en el principio de historicidad del método dialéctico de Richard Levins y Richard Lewontin:
«Cada problema tiene su historia en dos sentidos: la historia del objeto de estudio (…) y la historia del pensamiento científico acerca del problema, una historia dictada no por la naturaleza sino por las formas en que nuestras sociedades actúan y piensan sobre la naturaleza. Una vez que reconocemos que el “estado del arte” es un producto social, somos más libres para mirar críticamente la agenda de nuestra ciencia, su marco conceptual, y metodologías aceptadas, y para tomar decisiones de investigación conscientes» (Levins y Lewontin, 2007).
La cultura científica de la Guerra Fría
El evento que más influyó en la época de la posguerra en Estados Unidos, y en particular a la ciencia, fue la Guerra Fría (Bush, 1960): la guerra de Corea, la carrera armamentista de misiles y satélites, la competencia tecnológica con la Unión Soviética, la rápida aceleración de investigación espacial y el desarrollo de energía atómica. A partir de la segunda guerra mundial y durante la guerra fría la ciencia se vio ligada al proyecto militar: “La investigación básica es necesaria para la defensa nacional si los Estados Unidos no quieren encontrarse peleando la siguiente guerra con armas meramente mejoradas desde la pasada” (Bush, 1960). Así mismo también se conceptualizó la ciencia como un agente del progreso:
«El avance en la ciencia, al ponerse en uso práctico significará más trabajos, salarios más altos, horarios de trabajo más cortos, cosechas más abundantes, más tiempo libre para la recreación, para el estudio, para aprender cómo vivir sin el sofocante trabajo arduo que ha sido la carga común del hombre en épocas pasadas. Avances en la ciencia también traerán estándares de vida más altos, llevará a la prevención y cura de las enfermedades, promoverán la conservación de nuestros recursos nacionales limitados, y asegurarán los medios de defensa en contra de agresiones» (Bush, 1960, p. 10).
Y al servicio del capitalismo:
«No nos colocaremos a la delantera del comercio internacional a menos que ofrezcamos productos nuevos, más atractivos y baratos.
¿De dónde vendrán esos productos? ¿Cómo encontraremos maneras de hacer mejores productos a costos más bajos? La respuesta está clara. Debe haber una corriente de conocimiento científico nuevo para mover las ruedas del emprendimiento privado y público» (Bush, 1960, p. 18)
La cultura de la Guerra Fría entró en casi todas las disciplinas académicas y en particular las ciencias naturales y sociales (Kay, 2000). La entrada del discurso informacional a la biología coincide con ella. Antes de ello, durante la década de 1940, se hablaba de especificidad biológica. La especificidad biológica era la forma en la que los biólogos pensaban la manera de interactuar de las proteínas, enzimas, anticuerpos antes de la era de la información. Esta especificidad estaba determinada por la complementariedad espacial de las estructuras de las moléculas.
Después de la Segunda Guerra Mundial el gasto militar se volvió central en las políticas estadounidenses y fue la principal razón del “boom” económico. En 1949 el Departamento de Defensa utilizaba $13 billones de dólares anualmente, este presupuesto se elevó a $50 billones, y subió todavía más después del lanzamiento del Sputnik el 4 de octubre de 1957. En la década de 1950 el financiamiento de la ciencia provino principalmente del Departamento de Defensa, la NASA y la Comisión de Energía Atómica (AEC) que también estaba controlada por los militares. Este financiamiento se utilizó en la investigación y el desarrollo de armas nucleares, investigación espacial, computadoras electrónicas, guerra biológica, radiación y técnicas de control social (Kay, 2000). En cuanto a la biología, también se encontró en la década de 1950 atada al complejo militar, y en particular la genética. De hecho, en la primera mitad de la década, el Departamento de Defensa y la AEC componían más de la mitad del presupuesto federal para la biología y ciencias médicas. Además de becas y otras formas de financiamiento, también había laboratorios nacionales de biología molecular auspiciados militarmente por la AEC, por ejemplo: Argonne, Brookhaven y Oak Ridge. En particular la genética estuvo muy ligada a la AEC. En la década de 1950, proporcionó la mitad del financiamiento federal.
El financiamiento no fue externo a la ciencia, sino que tuvo influencia en la construcción del conocimiento. Como señala David Livingstone (Livingstone, 2003), el financiamiento influye profundamente en las formas de hacer ciencia. Por ejemplo, en las cortes italianas donde se valoraba el espectáculo y la extravagancia, la ciencia se practicaba de manera diferente respecto a Inglaterra, donde la credibilidad dependía de la sobriedad y los modales asociados con la nobleza. Las relaciones sociales, políticas y económicas de la ciencia afectan la cultura científica, la cual genera “diferencias cruciales entre lo que los estudiosos de la naturaleza investigan, quién tiene la posición para generar conocimiento, y qué intereses avanzan los proyectos científicos”(Livingstone, 2003). De esta forma, las ligas con el aparato militar de la ciencia de la década de 1950 no se limitaban al financiamiento. Los investigadores tenían que someterse a los procedimientos de seguridad militares y firmar un juramento de lealtad. La Guerra Fría alteró también los marcos institucionales, las estructuras organizacionales, y de manera importante los imaginarios y discursos científicos que trasminaron de los imaginarios y discursos culturalmente hegemónicos (Kay, 2000). Las tecnologías de la comunicación se volvieron esenciales para la guerra, entre ellas, los códigos y la transmisión de información. En este contexto, el problema de la especificidad biológica fue reformulado en términos de los discursos de la guerra fría: teoría de la información y la criptología (6).
El código genético no es un código
La historia de lo que se ha llamado el “código” genético y los esfuerzos para “romperlo” suele dividirse en dos fases, la formalista y la material. En la primera fase el problema “de la codificación” atrajo a físicos, matemáticos, criptólogos, analistas de sistemas y científicos de la computación. El primero de ellos fue George Gamow, quien fundó el RNA Tie Club, para tratar de “romper el código genético”. Este club se componía de 20 miembros, uno por cada aminoácido, muchos de los cuales, significativamente, laboraban al centro de proyectos de defensa.
“Gamow no sólo definió, articuló y trató de resolver el problema del código, sino que también trajo la poderosa cultura de la física de la posguerra, con sus múltiples vínculos militares, a recaer sobre las representaciones de la herencia y la vida. Y aunque su participación en la biología molecular fue temporal, dejó un legado duradero: su enfoque proporcionó el potente imaginario y el discurso con el que se constituyó ese objeto mítico -el código genético-” (Kay, 2000).
De acuerdo con la historiadora de la ciencia Lily Kay, un código es una manipulación sistemática del lenguaje (Kay, 2000), reglas de transformación lingüística (Kay, 1998). Como ya vimos, las metáforas textuales entraron a la biología por la coyuntura histórica en la que se vio ligada con la Guerra Fría. Una actividad central en la guerra y que significó un camino por el cual se desarrollaron las ciencias físicas y computacionales era la creación y desciframiento de códigos. La criptología fue una de las ciencias que más se incentivó durante la guerra. De esta manera, Gamow replanteó el problema de la síntesis proteica en términos de “romper el código enemigo”, así incidiendo en que “la vida se re-representara dentro de los regímenes de significación de la imaginación militar de la Guerra Fría”(Kay, 2000). Sin embargo, en los propios términos de la criptología el llamado código genético no es realmente un código, y por esta razón sus métodos fallaron y con ellos falló la primera fase en el estudio del “código genético”, la fase formal.
Lo que sucede es que todos los idiomas tienen restricciones lingüísticas en cuanto a las letras que pueden seguirse unas a otras en las palabras y frases, por ejemplo, en español no hay “n” antes de “p”, “m” antes de “t”, ni dos “o” juntas. Además, la frecuencia con la que letras específicas se siguen a otras es diferente en cada idioma, por ejemplo, en inglés es muy probable encontrar h después de t, y no así en español. Sin embargo, en el DNA no existen este tipo de restricciones ni probabilidades diferentes. Cuando los criptólogos corren sus programas de criptoanálisis estos lo que hacen es analizar las frecuencias de las letras vecinas y eso les da pistas para decodificar los textos. Los resultados son distribuciones que nunca siguen la distribución de Poisson, sin embargo, al correrlo en las secuencias genéticas obtenidas experimentalmente, la distribución seguía la de Poisson, es decir, una distribución aleatoria. En 1955 Gamow escribe:
“Distribuciones de frecuencia de vecinos se han listado para diversos idiomas y se utilizan extensamente en varias pruebas criptográficas. Sin embargo, en ninguna lengua escrita la distribución de letras vecinas sigue la de Poisson. (…) La ocurrencia de una distribución de Poisson es desmoralizante relativa a la posibilidad de descubrir un esquema de decodificación” (Gamow, Rich & Ycas, 1956).
Kay señala que la razón por la cual los análisis criptológicos no funcionaron en el DNA es porque éste no puede realmente conceptualizarse como un lenguaje, y los códigos corresponden al lenguaje. Sin embargo, Gamow y sus colaboradores, en lugar de reconocer el error en sus fundamentos conceptuales, lo que hicieron fue darle un significado biológico a su falta de resultados:
“Sin embargo, puede ser que el verdadero significado de lo anterior yace en una consideración de qué tanta ‘información’ puede transmitir una secuencia polipeptídica dada” (Gamow et al., 1956).
¿Cómo entró la “información” a la biología?
El término “información tiene dos significados muy diferentes. En el habla común la información se relaciona con “significado”. De acuerdo con Kay, “desde el s. XIV la palabra información ha significado la acción de informar: formación o moldeado de la mente y el carácter, entrenamiento, instrucción (…), conocimiento comunicado, noticias e inteligencia (en contraste con los datos)”(Kay, 2000). El diccionario de la Real Academia Española define “informar” como: “Enterar o dar noticia de algo” (7) e “información” como “Comunicación o adquisición de conocimientos que permiten ampliar o precisar los que se poseen sobre una materia determinada” (8), vinculando así el término “información con “noticia” y “conocimiento”. Así mismo, la enciclopedia colaborativa Wikipedia señala como características de la información: “significado”, “importancia”, “vigencia”, “validez” y “valor” (9).
Por otro lado, en la teoría matemática de la información de Claude Shannon (C. E. Shannon, 1948; Claude E. Shannon & Weaver, 1964), el término información tiene un significado técnico, en esta teoría la “información” es pura sintaxis, sin semántica, es decir sin significado. De acuerdo con Shannon:
“El problema fundamental de la comunicación es el de reproducir en un punto exacta o aproximadamente un mensaje seleccionado en otro punto. Frecuentemente, los mensajes tienen significado; esto es que se refieren o están correlacionados con algún sistema con ciertas entidades físicas o conceptuales. Estos aspectos semánticos de la comunicación son irrelevantes para el problema de ingeniería” (C. E. Shannon, 1948).
En la teoría de Shannon la información representa un parámetro cuantitativo cuya unidad es el “bit”. Y cuya fórmula es la siguiente:
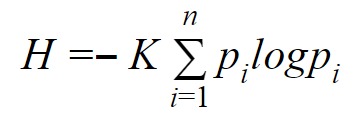
La cantidad de información no depende del significado, de hecho, un mensaje lleno de significado puede tener la misma cantidad de información que uno completamente sin sentido (Shannon & Weaver, 1964). La cantidad de información depende de la incertidumbre. Shannon conceptualiza la información como un mensaje seleccionado a partir de un conjunto de mensajes posibles utilizando el mismo conjunto de símbolos. La cantidad de información es mayor cuando las probabilidades de que cada símbolo sea seleccionado son las mismas ya que la incertidumbre que se resuelve con el mensaje es mayor. Si la probabilidad de que cierto mensaje sea seleccionado del conjunto de mensajes posibles es muy alta, entonces realmente el mensaje no contiene mucha información (Khan Academy, 2020; Shannon & Weaver, 1964).
Esta segunda definición de información fue por la que el término entró a la biología. Henry Quastler fue uno de los científicos que más prominentemente trabajó para aplicar la teoría de Shannon a la biología y por redefinir la especificidad biológica y química en términos informacionales. Él consideraba que, provisto que se definiera adecuadamente, la información podía proveer una medida exacta de la especificidad biológica. Quastler fue un médico originario de Viena con formación también en física, química y matemáticas. Llegó a estados unidos en 1932 refugiándose del fascismo europeo gracias al apoyo de la fundación Rockefeller. Formó parte del conjunto de científicos asociados a laboratorios y proyectos militares, trabajo en los laboratorios Argonne y Brookhaven, ambos de los cuales se originaron en el Proyecto Manhattan. Así mismo, también formó parte de los científicos que, fascinados por la teoría del control y de la información, buscaron articular una visión cibernética de la vida (Kay, 2000). Quastler se apegó al sentido matemático de la palabra información, buscando establecer un marco para cuantificar el flujo de energía en sistemas biológicos. Estableció varias colaboraciones con este propósito y llegó a tener cierto renombre en el medio. En este proceso Quastler y sus colaboradores introdujeron el vocabulario informacional a la biología, cómo se ejemplifica en las siguientes citas: “Una cinta de instrucciones lineal. Todo el hilo constituye el ‘mensaje.’ Este mensaje se puede romper en subunidades que pueden llamarse ‘párrafos’, ‘palabras’, etc.” (10) Y “Entonces la molécula de la proteína puede verse como el mensaje y los residuos de aminoácidos como el alfabeto” (11).
Organizó dos ediciones de un simposio llamado Information Theory in Biology. El primero, en 1952 es considerado por Kay como el “primer esfuerzo auténtico por reescribir la biología como una ciencia de la información” (Kay, 2000). Sin embargo, para la segunda edición en 1956 la posibilidad de aplicar la teoría de Shannon a la biología estaba puesta en duda, ya que todas las investigaciones arrojaban resultados negativos y que no eran concluyentes. En la publicación resultante del simposio Quastler escribe:
“La teoría de la información es muy fuerte en su lado negativo, es decir, en demostrar lo que no puede hacerse; en el lado positivo su aplicación al estudio de los organismos vivos no ha producido muchos resultados hasta ahora; no ha resultado en el descubrimiento de nuevos hechos, ni han sido probadas sus aplicaciones a hechos conocidos en experimentos críticos. Hasta la fecha, un juicio válido y definitivo al valor de la teoría de la información en la biología no es posible.” (Quastler, 1956)
Y el escepticismo del propio Shannon sobre la aplicabilidad de su teoría fuera de la ingeniaría también crecía (Kay, 2000). Sin embargo, a pesar del fracaso en desarrollar un marco sólido para la aplicación de la teoría de la información a la biología, el vocabulario informacional permaneció.
Simetría informacional en el desarrollo
Por otro lado, Paul Griffiths defiende la aplicación del concepto matemático de información en biología, pero señala que esa definición de información aplica igualmente a secuencias codificantes (12) que a muchos otros factores que no son genéticos, tanto internos como externos al organismo (Griffiths, 2017). Griffiths retoma la caracterización que Crick hace de la información dentro del marco de la teoría de Shannon que es: determinación específica de la secuencia de RNA por el DNA. Y observa que no sólo el DNA codificante (13) determina de manera específica la secuencia del RNA, sino también secuencias no codificantes y marcas epigenéticas que regulan splicing alternativo (14) y también señales ambientales a través de marcas epigenéticas (15). Así mismo, también muestra que no sólo la secuencia del RNA puede ser determinada específicamente, sino también otros aspectos en el desarrollo. El discurso dominante en la biología molecular posterior a los años cincuenta señaló a los ácidos nucleicos como los únicos que llevan información, y así han permanecido. Sin embargo, en una aplicación del término información congruente con la teoría de Shannon, no sólo los ácidos nucleicos llevarían información. En efecto, así es como lo trabajó Quastler, pero parecería que, en el proceso de metaforización, la información pasó a residir únicamente en los ácidos nucleicos.
Susan Oyama, Griffiths y colaboradores, denuncian la visión del desarrollo que divide los factores causales y procesos entre, por un lado, los genes y por el otro todo lo demás:
“La división entre rasgos innatos y adquiridos ha sido reemplazada por intentos de determinar la influencia relativa de los factores genéticos y ambientales en cada rasgo. La idea de un resultado especificado genéticamente ha sido reemplazada por un plan (16) genético y después por un programa genético. Pero todas estas explicaciones suponen que la clave para entender el desarrollo es entendiendo la interacción entre dos clases de recursos del desarrollo—genes y el resto. Todas son explicaciones dicotómicas del desarrollo.
La teoría de sistemas en desarrollo rechaza el enfoque dicotómico hacia el desarrollo: Los genes son sólo un recurso que está disponible en el desarrollo. Hay una simetría fundamental entre el papel de los genes y el del citoplasma materno o la exposición al lenguaje durante la infancia.” (Griffiths & Gray, 1994)
Que la metáfora informacional juega un papel central en esa visión dicotómica del desarrollo es palpable en una de las objeciones comunes a la relevancia causal de la epigenética en el desarrollo y la evolución:
“El problema con este tipo de sutilezas es que en última instancia la información extra (ej. metilación) (17) es provista por enzimas (metilasas) (18) codificadas en el genoma. La epigenética, per se, no agrega ninguna información nueva. Es sólo una consecuencia o un resultado de la información que ya está en el DNA (19).”
Esta crítica parece suponer que el hecho de que la estructura primaria (20) de una metilasa pueda ser especificada por el DNA (obviando el splicing alternativo), significa que la dinámica de metilación: qué segmentos se metilan, cuándo, en respuesta a qué, y demás procesos, también está contenida en el DNA. Este tipo de aseveraciones son claramente falsas, pero son facilitadas por el misticismo inherente a la mitología informacional al confundirse con significado.
Conclusión
Los términos como información, código, mensaje, alfabeto, texto y la visualización del DNA como un lenguaje provienen de la cultura científica de la Guerra Fría, en donde mucho del quehacer científico estaba ligado a las prácticas e intereses militares. En ese momento se desarrollaron tecnologías de la comunicación y control, la cibernética, la teoría matemática de la información de Shannon y la criptología. El trabajo de físicos, matemáticos, y otros científicos que participaban en proyectos de defensa, influyó para que la vida y la herencia se re-representaran en términos de los imaginarios y discursos de la guerra. No obstante, el DNA no es un lenguaje, ni un código, y es dudoso si puede generarse una teoría biológica de la información. De acuerdo con el marco teórico mediante el cual el término información entró a la biología, este no conlleva un significado. Sin embargo, el término fue perdiendo su definición original y se convirtió en una metáfora.
Al metaforizarse, la información pasó a concebirse como únicamente perteneciente a los ácidos nucleicos en la corriente biológica dominante. El retroceder sobre los pasos de la entrada de la información a la biología nos permite ver cómo el DNA no es por naturaleza información, sino de una manera socialmente construida y muy especializada. Esta asimetría sustenta una visión dicotómica del desarrollo. Apoya una concepción del desarrollo como la ejecución de un programa interno contenido en forma de información en el DNA. Una falsa dicotomía entre organismo y ambiente, en la cual el organismo está preespecificado internamente antes de su existencia en el mundo y sus múltiples relaciones con el ambiente.
Notas
1 DNA (ADN, en español): Es la molécula presente en todas las células, compuesta por secuencias de bases cuyo orden se puede traducir mediante complejos procesos celulares, en el orden de los aminoácidos en las proteínas.
2 Uno de los científicos premiados con el Nobel por el descubrimiento de la estructura de la molécula del DNA (o ADN).
3 Teleología: Que tiene una finalidad o propósito
4 El desarrollo u ontogenia en biología se refiere a todos los eventos y procesos que van desde un óvulo fecundado a un organismo adulto, y hasta su vejez y muerte
5 Genoma: Todo el ADN en los cromosomas de un organismo.
6 Criptología: estudio de los códigos
8 https://dle.rae.es/informaci%C3%B3n?m=form
9 https://es.wikipedia.org/wiki/Informaci%C3%B3n
10 Quastler y Dancoff citados por (Kay, 2000)
11 Herman Branson citado por (Kay, 2000)
12 Secuencia codificante se refiere a la secuencia de ADN que se traduce a un producto funcional, generalmente una proteína
13 Secuencia codificante de ADN
14 El splicing alternativo es una forma en la que las moléculas de ARN que se transcriben a partir de un gen en el ADN pueden procesarse de diferentes maneras dando como resultado secuencias diferentes de ARN que posteriormente resultan en secuencias diferentes de aminoácidos.
15 Las marcas epigenéticas son modificaciones químicas a la molécula del DNA u otras moléculas aledañas que modifican la expresión del gen, sin modificar la secuencia de bases en el DNA.
16 “bluebrint”: plano arquitectónico o diseño de ingeniería
17 La marca epigenética más estudiada
18 Enzimas que realizan la metilación
19 Larry Moran citado por Griffiths (2017)
20 La estructura primaria de una proteína, en este caso particular de la enzima metilasa, es la secuencia lineal de aminoácidos. Esta estructura todavía lleva a cabo diferentes pliegues y doblados, tomando una estructura tridimensional.
Referencias
Academy, K. (2020). La entropía de la información. Retrieved from https://es.khanacademy.org/computing/computer-science/informationtheory/moderninfotheory/v/information-entropy
Bush, V. (1960). Science the endless frontier. Washington DC: National Science Foundation.
Futuyma, D. J. (2017). Evolution (4th ed.). Sinauer Associates, Inc.
Gamow, G., Rich, A., & Ycas, M. (1956). The problem of information transfer from nucleic acids to proteins. In Advances in Biological and Medica Physics (pp. 23–62). NY: NY: Academic PRess.
Griffiths, P. (2017). Genetic, epigenetic, and exogenetic information. Interface Focus, 7(20160152), 1–8. https://doi.org/10.1098/rsfs.2016.0152 One
Griffiths, P., & Gray, R. D. (1994). Developmental Systems and Evolutionary Explanation. The Journal OfPhilosophy, 91(9), 277–304.
Kay, L. E. (1998). A Book of Life?: How the Genome Became an Information System and DNA a Language. Perspectives in Biology and Medicine, 41(4), 504–528. https://doi.org/10.1353/pbm.1998.0038
Kay, L. E. (2000). Who Wrote the Book of Life?: A History of the Genetic Code. Standford, California: Standford University Press.
Levins, R., & Lewontin, R. C. (2007). Biology under the influence.
Livingstone, D. N. (2003). Putting Sciencie In Its Place. Chicago: The university of Chicago Press.
Quastler, H. (1956). The status of information theory in biology. In Symposium on information theory in biology (pp. 429–432). NY: Pergamon Press.
Scott, J. W. (2009). Finding critical history. In J. M. Banner & J. R. Gillis (Eds.), Becoming historians (pp. 26–53). The university of Chicago Press.
Shannon, C. E. (1948). A Mathematical Theory of Communication. Bell System Technical Journal, 27(4), 623–656. https://doi.org/10.1002/j.1538-7305.1948.tb00917.xShannon, Claude E., & Weaver, W. (1964). The Mathematical Theory of Communication (1st ed.). Urbana: The university of Illinois Press.